
SciBase Journals
SciBase Oncology
ISSN 2691-7785
- Article Type: Research Article
- Volume 2, Issue 2
- Received: Aug 16, 2024
- Accepted: Oct 04, 2024
- Published Online: Oct 11, 2024
The Effects of Curcumin on ERα, p53, and p21 in the MCF-7 Breast Cancer Cell Line
Samantha Pfiffner1-3; Aisha Zanib1-3; Nina Prudhomme1-3; Kaylin Whittaker1-3; Raquel Yaldo1-3; Jehan Koujane1-3; Isabella Iskender1-3; Adele Salan1-3; Sumi Dinda1-3*
1Department of Clinical and Diagnostic Sciences, School of Health Sciences, Oakland University, Rochester, MI 48309, USA.
2Institute of Stem Cell and Regenerative Medicine, Oakland University, Rochester, MI 48309, USA.
3Center for Biomedical Research, Oakland University, Rochester, MI 48309, USA.
*Corresponding Author: Sumi Dinda
Department of Clinical and Diagnostic Sciences, School of Health Sciences, Oakland University, Rochester, MI 48309, USA.
Email: sdinda@oakland.edu
Abstract
Curcumin, a golden-yellow flavonoid derived from turmeric, has been used in traditional Chinese and Indian medicine for centuries due to its antioxidant and anti-inflammatory properties. Recent interest has grown in its potential therapeutic applications in cancer research, particularly breast cancer, which is the most commonly diagnosed cancer and the second leading cause of cancer death in women. This study investigates the effects of curcumin on the MCF-7 breast cancer cell line. MCF-7 cells were cultured in charcoal-stripped serum and treated with curcumin, alone and in combination with hormones and antihormones. After performing protein extraction and Bio-Rad Bradford protein assay, SDS-PAGE gel electrophoresis was performed followed by western blot analysis. Cellular viability was assessed using Propidium Iodine (PI) staining and confocal microscopy. Statistical analyses included Kruskal-Wallis testing and post-hoc Mann-Whitney U-tests, conducted using SPSS software. Western blot analysis revealed significant changes in ERα, p53, and p21 protein levels after 24-hour treatment with 5–100 μM curcumin. Specifically, curcumin induced an 80% reduction in ERα expression at 40 μM, and increased p53 and p21 levels by 40% and 80%, respectively. A 36% decrease in cellular viability was noted after six days of 40 μM curcumin treatment. Co-treatment with curcumin and E2 resulted in a 79% downregulation of ERα and increased p53 and p21 expression by 15% and 94%, respectively, compared to control. E2’s proliferative effect was mitigated by curcumin, with no significant changes in ERα cytolocalization observed. Curcumin shows potent anti-cancer properties in the MCF-7 breast cancer cell line, potentially contributing to understanding its molecular effects on breast cancer progression. This study supports further investigation into curcumin as a therapeutic agent in breast cancer treatment.
Keywords: Tumor suppressor; MCF-7; Antitumor agent; Natural anticancer; Cancer treatment.
Citation: Pfiffner S, Zanib A, Prudhomme N, Whittaker K, Dinda S, et al. The Effects of Curcumin on ERα, p53, and p21 in the MCF-7 Breast Cancer Cell Line. SciBase Oncol. 2024; 2(2): 1019.
Introduction
Breast cancer is the most commonly diagnosed cancer in women, impacting millions annually. With diverse risk factors and intricate molecular pathways involved in its progression, understanding the complexity of breast cancer is crucial for advancing prevention, early detection, and treatment strategies. In the year 2020, an estimated 2.3 million novel instances of breast cancer were diagnosed globally [1]. The mortality landscape of breast cancer continues to be a substantial global health challenge; in 2020, breast cancer led to approximately 42,170 deaths in the United States alone [2]. Estrogen, a primary sex hormone in females, plays a crucial role in the regulation of various physiological processes, including the development and maintenance of secondary sexual characteristics. In the context of breast tissue, estrogen exerts proliferative effects on epithelial cells, contributing to the intricate balance of cellular growth and differentiation. A comprehensive understanding of estrogen’s function in breast tissue is essential for unraveling its implications in both normal physiological states and pathological conditions such as breast cancer [3,4].
Estrogen has demonstrated in previous laboratory studies, as well as studies from our laboratory, proliferative effects on the hormone-dependent MCF-7 and T-47D breast cancer cell lines [3,5-8]. The primary receptor for E2 is estrogen receptor alpha, or ERα. In healthy breast epithelium, this receptor is present only 10% of the time, but when diseased breast epithelium is studied, it is shown to be present in approximately 50-80% of breast tumors. The presence of ERα significantly increases breast tumor mitotic activity in patients with mutated tumor-suppressor genes [9-10]. Furthermore, abnormal ERα expression has been shown to play a role in the development and progression of hormone-dependent breast cancer [10,11].
The MCF-7 breast cancer cell line used in this study was isolated from an infiltrating ductal carcinoma of the breast; the cell line is estrogen receptor positive and has the wild-type p53 gene [12]. The tumor suppressor protein p53 halts the cell cycle when DNA damage is found, induces apoptosis if damage is beyond repair, and ultimately protects cells from cancerous mutations. In approximately 50% of cancers, the TP53 gene is mutated, leading to unregulated cancer cell proliferation and progression [13]. The p21 protein is a cyclin-dependent kinase (cdk) inhibitor, which is induced by p53 in instances of DNA damage or oxidative stress [14,15]. As research advances, it seems that p21 may have a dual role as an oncogene and tumor suppressor; nevertheless, an upregulation of p21 has been shown to be synergistic with cancer treatment according to the literature [14,16]. Due to the intricate linking of these genes, proteins, and cancer progression, they are imperative to study in accordance with ERα to understand the progression of breast cancer.
Curcumin is a golden-yellow flavonoid compound derived from the turmeric (Curcuma longa) plant root which has been used in Chinese and Indian medicine for centuries. Its appeal to cancer research and therapeutics lies in its demonstrated antioxidant, anti-inflammatory, and anti-cancer properties [17-19]. Studies have shown curcumin to exhibit anticancer effects in Cancer Stem-Like Cells (CSCs), and also to restore expression of E-cadherin, a tumor suppressor, in breast CSCs [20,21]. Many other studies have also shown that curcumin suppresses breast cancer cells in vitro, leading to a concentration-dependent decrease in cancer cell growth, through a proposed mechanism of p53-induced apoptosis [22,23]. Our laboratory previously studied the effects of curcumin on the T-47D breast cancer cell line and found that curcumin significantly downregulated both p53 and ERɑ protein levels with a concomitant decrease in cell viability [24]. In this study, we investigate the effects of curcumin on ERɑ, p53, and p21 expression in the presence of hormones, and antihormones on the MCF-7 breast cancer cell line. This study will further contribute to the knowledge of curcumin’s anticancer properties and its actions on a molecular level in the proliferation and progression of breast cancer.
Materials and methods
Cell culture and treatment : The hormone-dependent MCF-7 breast cancer cell line (ATCC® HTB-133 TM; ATCC, Manassas, VA, USA) was regularly cultured at 37°C with 5% CO2, which mimics the ideal condition of the human body. The cells were cultured in Eagle’s minimum essential medium (Corning; Corning, NY, USA), which contained 1.5 g/L sodium bicarbonate, nonessential amino acids, l-glutamine, and sodium pyruvate. The cells were then plated in six-well plates (240,000 cells per well), and the medium was transitioned to a 5% dextran-coated charcoal-stripped fetal bovine serum (DCC-FBS; Santa Cruz Biotechnology; Dallas, TX, USA) (5% stripped serum) to avoid cellular exposure to endogenous steroids and growth factors within the FBS. This stripped serum allowed the cells to be at their basal metabolic rate at the time of treatment, confirming that any effects seen on the cells and the cells’ proteins are caused exclusively by the treatments they receive and not by any other components of the culturing media. The cells were cultured in this charcoal-stripped serum for 6 days. On the sixth day, the cells were treated with 2 μL of ligands for 24 hours. Control groups were treated with dimethyl sulfoxide (DMSO), the solvent used for curcumin treatments. Varying concentrations (5-100 μM) of curcumin were used in the concentration-dependent studies, while estrogens and antiestrogens were combined with 40 μM curcumin for the hormone studies. For estrogen treatments, 10 nM E2 was used. Additional treatments include a pure estrogen antagonist, 1 μM ICI (ICI 182, 780), and a selective estrogen receptor modulator, 1 μM TAM (Tamoxifen), was utilized.
Protein extraction and quantification
The cellular proteins were extracted with radioimmunoprecipitation assay buffer obtained from Santa Cruz Biotechnology (Dallas, TX, USA). The cells were then centrifuged at a speed of 15,000 ×g for 15 minutes at 4°C. After centrifugation, the protein supernatant was separated and used to prepare a protein assay based on the Bradford method (Bio-Rad; Hercules, CA, USA), which allowed for the quantification and normalization of the protein in each extracted sample by use of a spectrophotometer.
SDS-PAGE and western blot analyses
The extracted proteins were subjected to SDS-PAGE, prepared with SDS detergent and β-mercaptoethanol and followed by heating the supernatant for 3 minutes at 85°C. Equal amounts of protein were then loaded into a 7.5%–12.5% polyacrylamide gel and transferred to an Immobilon polyvinylidene fluoride membrane (Millipore; Bedford, MA, USA) by the process of electroblotting. To begin probing, 5% nonfat dry milk was used to block nonspecific proteins. The membranes were then probed with the corresponding primary and secondary antibodies for each protein of interest. For ERα, anti-ER anti-mouse monoclonal antibody (1:2000) from Santa Cruz Biotechnology (Santa Cruz, CA, USA) was used. To detect the primary antibody, a secondary goat anti-mouse IgG2a antibody (1:2000) was used. Anti-p53 anti-mouse monoclonal (1:2000) and anti-p21 BSA 5xTBS monoclonal (1:2000) primary antibodies were obtained from Cell Signaling Technology, Inc. (Danvers, MA, USA)and detected by secondary goat anti-rabbit antibodies (1:2000). All secondary antibodies were obtained from Jackson ImmunoResearch Laboratories, Inc. (West Grove, PA, USA). The specific band for each protein of interest was then visualized by the enhanced chemiluminescence method according to the instructions from Advansta (San Jose, CA. USA) and the Bio-Rad Chemi-Doc XRS + imaging system. The Western blots were subjected to quantification of the protein band density using the Image Studio Lite program, version 3.1 (LI-COR Biosciences; Lincoln, NE, USA).
Cell viability assay
Our cell viability assays demonstrate the number of live cells in a total population after treatments with ligands at varying concentrations for 6 days, with treatments administered every 2 days. The cell viability studies were cultured in 12-well plates (30,000 cells per well). Throughout the experiment, the medium was replenished every 48 hours. The plates were fed with 10% whole serum for the first 2 days to ensure confluency. The cells were then incubated in the presence of stripped serum for 6 days and quantified on the sixth day using the Cellometer Vision CBA Image Cytometry System software version 216 (Nexcelom Bioscience LLC.; Lawrence, MA, USA). This was accomplished via PI staining, which fluorescently tags dead cells. The number of dead cells versus total cells was quantified in order to calculate cell viability.
Immunofluorescence and confocal microscopy
Cells were plated onto coverslips in 12-well plates (30,000 cells per well) with the appropriate medium for the cell line. Subsequently, the cells were incubated with stripped serum for 6 days, and. immunolabeling was performed for ERɑ in MCF-7 cells. The distribution of three-dimensional fluorescent structures was analyzed using a Nikon Digital Eclipse C1-Plus confocal microscope (Nikon Instruments; Melville, NY, USA) and differential interference contrast (DIC) images were taken in parallel. NIS Elements AR software (Nikon Instruments) was used for image analysis.
Statistical analyses
The results are expressed as mean ± SEM. Statistical significance was determined by the Kruskal–Wallis test, followed by post hoc analysis using the Mann–Whitney U test. Differences are considered significant at P<0.05. In all figures: *p<0.05, **p<0.01, and ***p<0.001. Statistical analyses were carried out using SPSS for Windows version 11.5 (SPSS Inc.; Chicago, IL, USA).
Results
Concentration-dependent effects of curcumin on Erα, p53, and p21 protein expression
Figures 1A, 1B and 1C represent the results of the concentration-dependent effects of curcumin on ERα, p53, and p21 expression respectively. Western blot analysis revealed a concentration-dependent decrease in ERα protein expression with treatments of 5–100 μM curcumin. An 80% decrease in ERα expression was observed with treatment of 40 μM curcumin when compared to control (Figure 1A). A concentration-dependent increase in expression of both p53 and p21 was observed after treatment of 5–100 μM curcumin. A 130% increase in p53 protein levels was seen after treatment with 100 μM curcumin and compared to control (Figure 1B). Expression of p21 was increased by 80% when compared to control after treatment with 80 μM curcumin (Figure 1C).
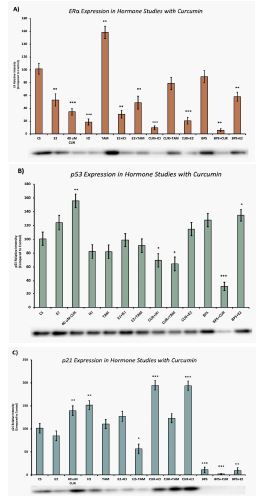
Effects of curcumin in combination with estrogens and antiestrogens on ERα, p53, and p21 protein expression
Figures 2A, 2B and 2C represent the effects of hormonal studies with curcumin on ERα, p53 and p21 expression, respectively. Treatments of E2 (10 nM) show an expected decrease in ERα expression in the MCF-7 cell line (Figure 2A). Similar effects are seen with ICI, as well as the combination of E2 and ICI. TAM showed an expected increase in ERα expression, which normalized compared to control with curcumin co-treatment. A synergistic effect was observed on the downregulation of ERα expression when curcumin (40 μM) was combined with E2 or ICI. An 80% decrease in ERα expression was observed following treatment of curcumin and E2. Combined treatments of curcumin and E2 showed no significant effect on p53 when compared to control (Figure 2B). However, when curcumin was combined with ICI, TAM, and Bisphenol S (BPS), significant reduction in p53 expression was observed when compared to control (Figure 2B). A maximum 60% reduction in p53 expression was noted when curcumin was combined with BPS. Treatment of curcumin and E2 resulted in a 94% increase in p21 when compared to the control (Figure 2C). Similarly, when curcumin was combined with ICI a 94% increase in p21 was also demonstrated when compared to the control.

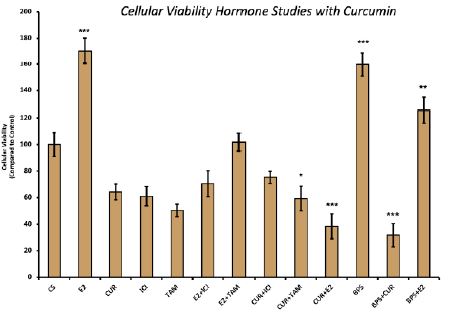
Effects of curcumin in combination with estrogens and antiestrogens on cell viability
Treatment with E2 displayed a proliferative effect on MCF-7 cell viability, which is to be expected. This effect of E2 was reversed when cells were treated with a combination of E2 and curcumin, which showed a 62% decrease in cellular proliferation when compared to control (Figure 3). Similarly, the proliferative effect of BPS on MCF-7 cells was reversed with co-treatment of BPS and curcumin, which demonstrated a 69% decrease when compared to the control.
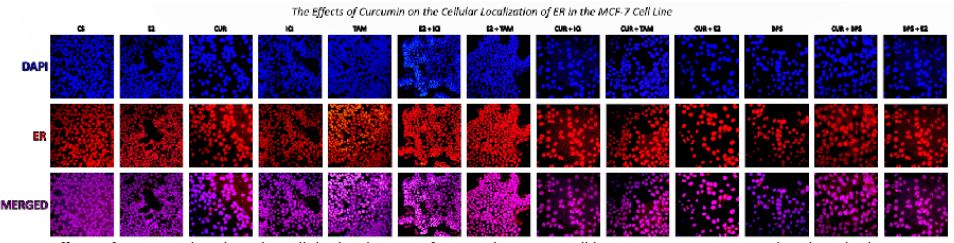
Effects of curcumin on the cellular localization of ERα
Confocal images show that ERα is localized in the control (DMSO) intranuclearly, as expected (Figure 4). Following treatment of cells with 10 nM E2, 1 μM ICI, 40 μM curcumin, TAM, BPS and combinations of these, there is no observed change in cellular localization of ERα.
Discussion
Breast cancer remains a pervasive global health challenge, with millions of cases diagnosed annually and significant mortality rates, emphasizing the urgency for enhanced prevention, early detection, and treatment strategies [1,2]. The pivotal role of estrogen, particularly through its proliferative effects on breast tissue, underscores the intricate balance in cellular growth and differentiation, with abnormal ERα expression implicated in hormone-dependent breast cancer development and progression [3,4]. Notably, the interplay of key regulators such as ERα, p53, and p21 in breast cancer pathogenesis is underscored, with mutations in these genes contributing to unregulated cell proliferation. Curcumin, derived from the turmeric plant, has garnered attention for its profound antioxidant, anti-inflammatory, and anti-cancer properties. Extensive research suggests curcumin’s potential to inhibit cancer cell growth, particularly in breast cancer, and modulate key molecular pathways involved in proliferation [17-21]. Previously, our laboratory studied the effects of curcumin on the T-47D breast cancer cell line and found curcumin to significantly downregulate both p53 and ERɑ protein levels with a concomitant decrease in cellular viability [24]. Our current studies expand on this investigation and contribute to the understanding of curcumin’s mechanism of action on the hormone-dependent breast cancer cell line, MCF-7.Overall, western blot analysis demonstrates a concentration-dependent decrease in ERɑ expression with increasing curcumin concentrations, depicting an 80% reduction in expression when the optimum concentration of 40 μm curcumin is compared to the control. This effect is maintained upon co-treatment of E2 and curcumin, which again demonstrates an 80% decrease in ERɑ expression when compared to the control. This is consistent with our findings in the cellular viability experiment, which depicts a corresponding 62% decrease in cellular viability following the same treatment combination. This may indicate the relationship between ERɑ expression and breast cancer proliferation. In addition, ERɑ expression is significantly decreased when curcumin is combined with the antiestrogen ICI, even more so than when either compound is used in isolation. In addition, there is no observed change in cellular localization of ERα following hormone treatment. These effects of curcumin on ERɑ expression are consistent with the results seen in our previous studies within the T-47D cell line [24].
The protective, anti-cancer effects of tumor suppressor gene TP53 have been well documented, along with the mechanism of its downstream effector protein p21 [13-15]. In our concentration studies, curcumin treatment demonstrates a concentration-dependent increase in both p53 and p21 levels when compared to the control, with a maximum 132% and 80% increase in each protein respectively. This increase in p21 expression was sensitive to the co-treatment of curcumin and E2, demonstrating a 94% increase when compared to the control. Additionally, when treated with combinations of curcumin and antiestrogens ICI and TAM, this downstream effect on p21 still prevails, with demonstrated 94% and 23% increase respectively. This portrays the continuous protective effects of curcumin even in the anti-estrogen setting. These results are corroborated by the corresponding decrease in cellular viability when curcumin is combined with antiestrogens ICI and TAM; cellular viability decreased by 25% and 41% after these respective treatments when compared to the control. Our results regarding the p53 studies in the MCF-7 cell line are in contrast to our findings previously in the T-47D cell line, which displayed a concentration- dependent decrease in p53 levels following curcumin treatment [24]. This contradiction is likely because the T-47D cell line harbors a TP53 gene mutation while the MCF-7 cell line’s TP53 gene is wild-type.
Moreover, estrogen treatment alone has been well documented to increase the proliferation of breast cancer cells, which is depicted in our cellular viability experiment by a 70% increase in viability following treatment of E2 when compared to the control [5-8]. However, when curcumin and E2 are combined as co-treatment, curcumin’s antiproliferative effects are able to suppress the proliferative effects of E2, which is depicted by the 62% decrease in cellular viability of this combination. This effect is even more dramatic than the decrease in cellular viability noted when cells are treated with TAM or ICI in isolation.
On a final note, our findings contribute to the current discussion of breast cancer treatment and prevention on the molecular level; They offer an improved understanding of the effects of curcumin not only on ERα, but also on p53 and p21 expression, and their corresponding effects on cellular viability. Our studies delineate the clear anti-estrogenic, and anti-cancer nature of curcumin, and corroborate its protective effects on an additional breast cancer cell line, MCF-7. The combination of decreased ERα expression, increased p53 and p21 expression, and decreased cellular viability following curcumin treatment is promising in terms of cancer treatment and/or prevention. Moreover, the fact that these effects are maintained in the setting of estrogenic and anti-estrogenic compounds is even more encouraging. Further studies are indicated to expand upon this research in the clinical setting.
Declarations
Conflicts of interest: The authors declare no conflicts of interest.
Ethical Approval: Not applicable.
Consent to Participate: Not applicable.
Consent to Publication: Informed consent to publication was obtained from relevant participants.
Availability of Data and Materials: The raw data supporting the conclusions of this manuscript will be made available by the authors, without undue reservation, to any qualified researcher.
Funding: Not applicable.
Acknowledgments: We would like to acknowledge Ms. Mia Morse for her technical assistance to this project.
References
- World Health Organization. Breast cancer: prevention and control. 2021. https://www.who.int/cancer/detection/breastcancer/en/
- American Cancer Society. Breast Cancer Facts & Figures. 2021. https://www.cancer.org/cancer/breast-cancer.html
- Jensen EV, Jordan VC. The estrogen receptor: a model for molecular medicine. Clinical Cancer Research. 2003; 9(6): 1980–1989. doi:10.1158/1078-0432.CCR-08-0784.
- Anderson E, Clarke RB. Steroid receptors and cell cycle in normal mammary epithelium. Journal of Mammary Gland Biology and Neoplasia. 2004; 9(1): 3–13. doi:10.1023/B:JOMG.0000048779.13361.8b
- Szmyd M. The effects of black cohosh on the regulation of estrogen receptor (ERα) and progesterone receptor (PR) in breast cancer cells. Breast Cancer. 2018; 10: 1–11.
- Siebert AE, Sanchez AL, Dinda S, Moudgil VK. Effects of estrogen metabolite 2-methoxyestradiol on tumor suppressor protein p53 and proliferation of breast cancer cells. Syst Biol Reprod Med. 2011; 57(6): 279–287.
- Dinda S, Sanchez A, Moudgil VK. Effects of LY117018 (a SERM analog of raloxifene) on tumor suppressor proteins and proliferation of breast cancer cells. Horm Mol Biol Clin Investig. 2010; 2(1): 211–217.
- Powell CE, Soto AM, Sonnenschein C. Identification and characterization of membrane estrogen receptor from MCF7 estrogen-target cells. J Steroid Biochem Mol Biol. 2001; 77(2–3): 97–108.
- Tung N, Wang Y, Collins LC, et al. Estrogen receptor positive breast cancers in BRCA1 mutation carriers: clinical risk factors and pathologic features. Breast Cancer Res. 2010; 12(1): R12.
- Jia M, Dahlman-Wright K, Gustafsson JÅ. Estrogen receptor alpha and beta in health and disease. Best Pract Res Clin Endocrinol Metab. 2015; 29(4): 557–568.
- Yip CH, Rhodes A. Estrogen and progesterone receptors in breast cancer. Future Oncol. 2014; 10(14): 2293–2301.
- Powell CE, Soto AM, Sonnenschein C. Identification and characterization of membrane estrogen receptor from MCF7 estrogen-target cells. J Steroid Biochem Mol Biol. 2001; 77(2–3): 97–108.
- Duffy MJ, Synnott NC, Crown J. Mutant p53 as a target for cancer treatment. Eur J Cancer. 2017; 83: 258–265. doi:10.1016/j.ejca.2017.06.023
- Shamloo B, Usluer S. p21 in cancer research. Cancers. 2019; 11(8): 1178. https://doi.org/10.3390/cancers11081178
- Abbas T, Dutta A. p21 in cancer: intricate networks and multiple activities. Nat Rev Cancer. 2009; 9: 400–414.
- Georgakilas AG, Martin OA, Bonner WM. p21: A two-faced genome guardian. Trends Mol Med. 2017; 23: 310–319.
- Choudhuri T, Pal S, Agwarwal ML, Das T, Sa G. Curcumin induces apoptosis in human breast cancer cells through p53-dependent Bax induction. FEBS Lett. 2002; 512(1–3): 334–340.
- Mudduluru G, George-William JN, Muppala S, et al. Curcumin regulates miR-21 expression and inhibits invasion and metastasis in colorectal cancer. Biosci Rep. 2011; 31(3): 185–197.
- Deguchi A. Curcumin targets in inflammation and cancer. Endocr Metab Immune Disord Drug Targets. 2015; 15(2): 88–96.
- Kim SR, Park HJ, Bae YH, et al. Curcumin down-regulates visfatin expression and inhibits breast cancer cell invasion. Endocrinology. 2012; 153(2): 554–563.
- Kim HI, Huang H, Cheepala S, Huang S, Chung J. Curcumin inhibition of integrin (α6β4)-dependent breast cancer cell motility and invasion. Cancer Prev Res. 2008; 1(5): 385–391.
- Nejati-Koshki K, Akbarzadeh A, Pourhassan-Moghaddam M. Curcumin inhibits leptin gene expression and secretion in breast cancer cells by estrogen receptors. Cancer Cell Int. 2014; 14: 66.
- Liontas A, Yeger H. Curcumin and resveratrol induce apoptosis and nuclear translocation and activation of p53 in human neuroblastoma. Anticancer Res. 2004; 24(2B): 987–998.
- Hallman K, Aleck K, Dwyer B, et al. The effects of turmeric (curcumin) on tumor suppressor protein (p53) and estrogen receptor (ERα) in breast cancer cells. Breast Cancer (Dove Med Press). 2017; 9: 153–161. doi:10.2147/BCTT.S125783