
SciBase Journals
SciBase Oncology
ISSN 2691-7785
- Article Type: Review Article
- Volume 3, Issue 1
- Received: Sep 12, 2025
- Accepted: Nov 14, 2025
- Published Online: Nov 21, 2025
Hippocampus and its significance in cranial irradiation
Bhuvana J*
Department of Radiation Oncology, Gujarat Cancer and Research Institute (GCRI), India.
*Corresponding Author: Bhuvana J
Department of Radiation Oncology, Gujarat Cancer and Research Institute (GCRI), Ahmedabad, Asarwa, New Civil Hospital Campus, Ahmedabad, Gujarat 380016, India.
Tel: 8248170875;
Email: Bhuvijayasree@gmail.com
Abstract
Purpose: Hippocampus is an intricate structure which plays a crucial role in neurocognitive function. It contributes primarily to explicit memory tasks, spatial memory attention, and problem-solving ability. Cranial irradiation, while a common modality of treatment in numerous cancers, is associated with brain parenchymal injury. Neural stem cell niches in the subgranular zone of hippocampal formation is particularly susceptible to radiation induced injury, which in turn is associated with deleterious effect on neurocognitive functions. A maximum dose of < 16Gy and D100 of < 9Gy is the usually preferred hippocampal constraints.
Conclusion: The cognitive preservation has more profound effect in paediatric and young population and therefore, anatomical avoidance of hippocampal formation while delivering cranial irradiation should be routinely practiced. It is necessary that our understanding of dose-dependent radiation-induced NSC dysfunction, the growing knowledge of NSC niches in the brain and development of high precision radiation delivery techniques, should guide us to design and execute NSC-preserving radiotherapy.
Citation: Bhuvana J. Hippocampus and its significance in cranial irradiation. SciBase Oncol. 2025; 3(1): 1022.
Introduction
The hippocampus is a complex structure involved in neurocognitive function of brain. Cranial radiotherapy has been used consistently either as a curative, palliative or prophylactic treatment modality in cancer management. Nevertheless, various studies have highlighted the deleterious effects of radiation on Neurocognitive Function (NCF), which is particularly detrimental to learning, memory and spatial processing. These studies provided convincing evidence of the association of NCF and neural stem cell niches in the subgranular zone of hippocampal formation. It is believed that radiation kindles damage to these stem cells which ultimately results in neurocognitive deficits in patients after cranial irradiation [1,2]. The interest in the hippocampus can be attributed to the fact that it can be clearly delineated on magnetic resonance imaging and, with advanced conformal radiotherapy techniques, the hippocampus can be safely avoided while delivering cranial radiation therapy [3].
History of hippocampus
In 1587, Julius Caesar Arantius, an Italian anatomist and surgeon, described a ridge running along the floor of the temporal horn of the lateral ventricle. He analogized this ridge first to the white silkworm and later to the seahorse. Over the course of time, this structure was assigned various names, like “pes hippocampi” (1672), “ram’s horn” (1732), “cornu ammonis” (1742), but the term “hippocampus” was dominantly used in the literature and eventually became the recognized term [4]. The term “hippocampus” is derived from the Latin word hippocampus which inturn finds its roots back to the Greek word hippokampus (hippos, meaning “horse,” and kampos, meaning “sea monster”),
The exact function of this complex structure eluded researchers for a long time. But in the 19th century, studies focussing on behavioral function of the hippocampus started gaining momentum. In 1878, Paul Broca described the limbic lobe and considered the hippocampus to be a part of it. The limbic system is generally involved in cognitive functions like spatial memory, learning, motivation, emotional processing, and social processing. Broca stated that this system is comprised of circumventricular structures like subcallosal, cingulate, parahippocampal gyri and infolded hippocampal formation. In the following years, structures included under the “limbic system” have escalated and have become the fodder for various debates regarding the relevance of this term either as a neuroanatomical or functional entity [5].
Papez, in 1937, published a landmark paper entitled, “A proposed mechanism of emotion” [5]. Papez proposed that the hypothalamus, the anterior thalamic nuclei, the gyrus cinguli, the hippocampus, and their interconnections form Papez’s circuit, which participate in the functions of central emotion. He considered hippocampus to be a central component of emotional expression which collects and channels the perceptions of emotional situations to the hypothalamus [5].
The behavioral function of hippocampus was further supported by Mclean in 1954 [6]. McLean proposed the model of the Triune Brain, which consists of the reptilian complex or the R-complex, the paleomammalian complex or the limbic system, and the neocortex. The limbic system consists of the septum, amygdala, hypothalamus, hippocampal complex, and the cingulate cortex. He claimed the limbic system to be responsible for motivation and emotion related to sexuality and reproductive behavior, feeding, and parental practice [7]. Hippocampal formation was also associated by early scientists solely with olfaction. However, in 1947, Alf Brodal concluded that the hippocampus could not function only as an olfactory structure because anosmic mammals such as dolphins had a substantial hippocampal formation [8].
Hippocampus was subsequently associated with response inhibition models. Douglas in 1967 demonstrated that hippocampal lesions produce an inability to withhold a response [9]. In the following year, Kimble presented that the mammalian hippocampus is important in behavioral situations in which Pavlovian internal inhibition occurs [10]. Similar to these concepts of “response inhibition” suggested by Douglas or “internal inhibition” favored by Kimble, Altman et al. hypothesized hippocampal function akin to powerful brakes applied in the normal animal [11].
The role of the hippocampus in memory was highlighted by Scoville & Milner in 1957. Bilateral Medial Temporal-lobe resection with damage to anterior hippocampus and Hippocampal gyrus in psychotic patients and patients with intractable seizures, leads to persistent impairment of recent anterograde & retrograde memories, but remote memories were intact. There was no effect on patients’ integral personality or performing complex tasks [12].
Another most appreciated model is spatial theory. O’Keefe and Nadel claimed hippocampus mediates a neural representation of physical space, otherwise known as a cognitive map [13]. In addition, O’Keefe and Nadel proposed the firing rate of some hippocampal neurons in rats increased based on the surrounding environment [14]. This lead to the belief that spatial information is encoded within hippocampal structures, and it is pertinent for spatial learning and memory. Thus, the contributions of various researchers lead to better understanding of the anatomy and functioning of the hippocampus.
Anatomy of hippocampus
The hippocampus is a protuberance in the floor of the temporal horn of the human lateral ventricle. It is a component of the limbic circuit, which includes white matter tracts such as the fimbriae and fornices, and gray matter structures such as the amygdala and para hippocampal gyrus. The term hippocampal formation includes complex cytoarchitecture comprising distinct adjoining regions including the hippocampus, subiculum, presubiculum, parasubiculum, and entorhinal cortex [4,15]. Hippocampal formation consists of two to five digitations which form the pes hippocampi [16]. The adjective hippocampal, as in “hippocampal lesions”, refers to regions beyond these cytoarchitectonic regions.
The hippocampus consists of two U-shaped interlocking laminae: the cornu ammonus and the dentate gyrus. Celebrated neuroanatomist Lorente de Nó in 1934 classified Ammon’s Horn into 3 histological subdivisions, which comprise CA3, CA2, and CA1, where CA is derived from cornu ammonis [17]. The Dentate Gyrus (DG) is a trilaminate cortical region with a characteristic V or U shape. The hippocampal fissure separates CA3 and CA1 fields in DG. The granule cell layer located between these two fields is called the Suprapyramidal blade, and the region opposite to this is the Infrapyramidal blade. The bridge between these two blades is the crest.
The dentate gyrus comprises three layers. The superficial layer is a relatively cell-free, called the molecular layer and deep to this layer is the granule cell layer, which is a densely packed layer of four to eight granule cells in thickness. The granule cell and molecular layers form a V- or U-shaped structure that encloses the third layer, the polymorphic cell layer. The polymorphic layer forms a narrowband called the hilus [15].
The Hippocampus consists of a pyramidal cell layer which forms the principal cellular layer. It is tightly packed in CA1 and loosely packed in CA2 and CA3. Hippocampus comprises a cell-free layer called the stratum oriens, a narrow acellular zone called the stratum lucidum, a suprapyramidal region called stratum radiatum and stratum lacunosum-moleculare, in which fibers from the entorhinal cortex terminate. The major source of hippocampal input is the hippocampus itself, as most of its synaptic input arises within its own boundaries [15].
Majority of sensory information reaching the hippocampus enters through the Entorhinal Cortex (EC) and after processing, gets relayed back to the neocortex. The laminar organization of EC comprises 6 layers, of which II, III, V, VI are cellular and I and IV are plexiform layers but lack a granular cell layer [18]. EC divides into medial and lateral components. Medial EC interacts with cortical areas that are involved in spatial processing, while lateral EC is connected with those areas involved in processing of object information, attention and motivation [19].
Subiculum is one among the primary output structures of hippocampal formation. Subiculum projects to Presubiculum and parasubiculum. The presubiculum comprises Brodmann’s area 27. The parasubiculum comprises the Brodmann’s area 49 [20]. Psalterium is the commissural fiber that extends across the midline to form the hippocampal commissure [15].
Radiologically, the hippocampus appears hypointense on T1 weighted MRI images (Figure 1). It appears as a curved banana shaped structure in axial MRI sequences. The hippocampus remains medial to the temporal horn of the lateral ventricle throughout its extent. The anterior boundary of the hippocampus is defined by the anterior edge of the temporal horn. The medial boundary of the hippocampus is defined by the “boomerang-shaped” uncus and posterocranially quadrageminal cistern.
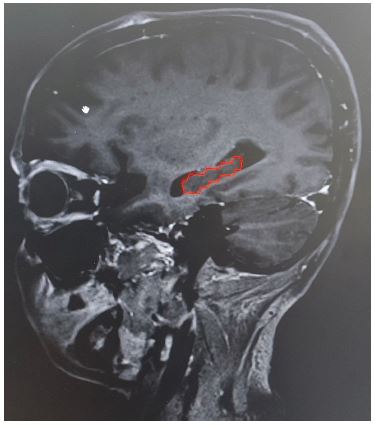
Hippocampal formation is distinct from neocortical areas as it lacks reciprocal connections. Majority of neocortical input reaching the hippocampal formation go through the EC. Superficial layers of the EC comprise axonal projections to the DG, which forms the major hippocampal input pathway called the perforant path. This pathway is nonreciprocal, as the dentate gyrus does not project back to the entorhinal cortex [21,18]. Also, there is almost no commissural interaction between the hippocampal formations located on each side of the human brain [15].
Major fiber systems associated with the hippocampal formation are the angular bundle, the fimbria-fornix pathway and dorsal and ventral commissures. Angular bundle carries fibers between the EC and the other fields of the hippocampal formation. The Fimbria-fornix pathway interconnects the hippocampal formation with the basal forebrain, hypothalamus, and brain stem regions. The dorsal and ventral commissures connect the hippocampal formation of one hemisphere with the contralateral hemisphere [15].
Adult neurogenesis
Over 5 decades ago, Joseph Altman and colleagues provided the initial evidence of adult neurogenesis occurring in an intact mammalian brain involving regions like the dentate gyrus and the olfactory bulb, and these findings were corroborated by others [22,23]. Neural Stem Cells (NSC) are quiescent with cell-cycle time of 28 days and generate transiently dividing progenitor cells that are characterized by a cell-cycle time of 12 h [24]. Progenitor cells located in the dentate gyrus give rise to new cells in the dentate gyrus itself. These cells are primarily concentrated in the sub granular zone which is located between the granule cell layer and the hilus. The sub granular zone contains sporadic collection of progenitor cell clusters which produce some daughter cells that retain the ability to divide and the remaining undividable cells become either neurons or glia. These new cells migrate to the granule cell layer and differentiate into neurons.
Functions of hippocampus
The hippocampal formation consists of highly intrinsic neuronal networks that integrate highly processed, multimodal sensory information from various neocortical sources. The principal role of the hippocampus in memory function was first identified in a patient H.M., who underwent a bilateral medial temporal lobectomy for the relief of medically intractable epilepsy. Following the procedure, H.M. showed a severe anterograde amnesia characterized by impairment in declarative memory but the remaining components of his neurocognition, including perception, intelligence, working memory, and motor learning skills remained largely intact. It was concluded that hippocampal injury results in impairment of declarative memory regardless of the sensory modality in which information is presented, without affecting perception and intellectual functions [25].
Memory function has been associated with the pyramidal and granule cells located in the dentate gyrus of the hippocampus. Explicit or declarative memory refers to the conscious recollection of prior experiences. Implicit or nondeclarative memory refers to changes in performance or behavior, produced by prior experiences, and does not require any conscious recollection of those experiences. Hippocampus plays a role in explicit memory tasks, and implicit memory tasks are usually spared in hippocampal lesions, although they may be used in implicit tasks that evoke declarative processes [15,26]. Hippocampus is required for declarative memory retrieval and, to some extent, encoding. In amnestic syndrome, central impairment is one of retrieval rather than encoding or storage. But studies have shown that encoding & retrieval processes are not unique to the hippocampus, but appear to be present throughout the median temporal lobe [27,28].
The visual-paired comparison task, despite its implicit nature, is dependent on the hippocampal region and the adjacent medial temporal lobe cortices [26]. Also, hippocampal damage leads to temporal gradients i.e., retrograde amnesia marked by greater loss of memory for occurrences from the recent past than for occurrences from long ago [29]. The differentiation of function between the hippocampal region and the adjacent cortical structures like the parahippocampal gyrus is not currently apparent, and it can be presumed that the latter support associative, recollective, or source components of declarative memory [15].
Spatial memory is an example of complex, declarative memory. Hippocampus plays a major role in performing spatial memory tasks, but its function is more mnemonic and is not involved in non-mnemonic aspects of cognition, including spatial processing. Therefore, complete hippocampal loss results in inability to acquire new spatial information but does not affect spatial processing [15]. Broadbent & Squire reflected that spatial memory and object recognition memory are affected by Hippocampal lesions. They concluded that spatial memory is more vulnerable to hippocampal dysfunction as spatial memory performance requires more hippocampal tissue than does recognition performance [17].
The place cell of hippocampus codes for the current location in a spatial environment. These cells code for places, directions, and speed of movement and are present in the hippocampus proper and subicular complex [30].
Radiation induced brain injury
Radiation Therapy Oncology Group (RTOG) describes late toxicity as adverse effects that occur after 90 days of commencement of treatment. The common late effects of cranial Radiotherapy (RT) include neurocognitive deficits. They are usually not self-limited and may have severe consequences. Leukoencephalopathy and radiation necrosis are other potential late toxicities associated with cranial RT [31].
Armstrong et al. proposed a biphasic pattern of memory deficits following Whole Brain Radiotherapy (WBRT). Transient decline in mental functioning was observed at 4 months after WBRT, followed by an improvement in neurocognitive functioning, and then an irreversible impairment month to years later [32]. Tallet et al. Performed a meta-analysis to assess the NCF after WBRT at various prophylactic doses in patients with brain metastases and found a decline of 31-57% in NCF at 3 months, and 48-89% at 1 year, emphasizing the belief that WBRT causes neurocognitive decline [33].
The mechanisms inducing this neurocognitive decline are varied. Radiation results in injury to brain parenchyma. Initially, radiation-induced brain injury was considered as the result of DNA damage. But preclinical studies conducted in the last two decades demonstrate that radiation-induced late effects, including cognitive impairment, are hypothesized to occur due to dynamic interactions between radiation and multiple cell types within the brain, including astrocytes, endothelial cells, microglia, neurons and oligodendrocytes [34].
Radiation induced necrosis was extensively studied by Greene-Schloesser et al. Damage to vascular and glial cells, reductions in hippocampal neurogenesis, impairment of synaptic plasticity and chronic inflammation were considered as the major mechanisms resulting in brain injury [35].
Damage to vascular and glial cells is an acute process. WBRT leads to time- and dose-dependent reductions in the brain endothelial cells. Acute increase in capillary rarefaction and tissue hypoxia is observed in the hippocampus after cranial irradiation. Radiation-induced loss of oligodendrocyte type-2 astrocyte (O-2A) progenitor cells results in a failure to replace oligodendrocytes, ultimately resulting in demyelination and white matter necrosis.
Reductions in hippocampal neurogenesis are associated with radiation-induced cognitive impairment. Cranial irradiation can lead to a dose-dependent decrease in NSCs, decreased proliferation of surviving NSC, and decreased differentiation of NSC into neurons.
Irradiating the brain elicits an inhibition of hippocampal Long-Term Potentiation (LTP). Homer1a is a synaptic plasticity early response gene essential for the activity-dependent regulation of excitatory synaptic transmission. Homer1a exhibited decreased expression in both the hippocampus and cortex 2 months after WBRT, resulting in impaired synaptic plasticity.
In response to cranial irradiation, a chronic inflammatory process occurs in the brain which includes elevation of inflammatory cytokines in the brain up to 6 months post irradiation, a marked increase in the number of activated microglia in the neurogenic zone of the dentate gyrus, increased expression of the CCR2 receptor in the sub granular zone 9 months post irradiation and persistent microglial and astrocyte activation. This results in radiation-induced brain injury, including cognitive impairment.
Rationale behind hippocampal avoidance
Jalali et al., evaluated Intelligence Quotient (IQ) scores in patients treated with radiotherapy for benign and low-grade brain tumors and observed a significant correlation association between IQ decline and dose to the left temporal lobe [36]. But this anatomic explanation was challenged in favour of the “stem-cell compartmental” hypothesis. The discovery of persistent neurogenesis in the Sub Ventricular Zone (SVZ) of the lateral ventricles and the Sub Granular Zone (SGZ) of the hippocampal dentate gyrus in adults opened a new arena of NCF research [37].
The hallmarks of radiation-induced cognitive impairment are decrements in verbal memory, spatial memory, attention, and problem-solving ability, with incidence and severity increasing over time [34,38]. As previously discussed, the hippocampus plays a critical role in these functions and therefore, we can safely postulate radiation induced hippocampal injury as the cause of these cognitive deficits.
A modest dose of radiation results in increased rates of hippocampal apoptosis, decreased rates of proliferation, and a decrease in adult neurogenesis, which is in turn associated with cognitive deficits. There is no significant apoptosis in other areas of the cerebrum, and no loss of function is observed in hippocampal-independent tasks [39].
This was also supported by Monje et al., who observed that after a single fraction of 10Gy to the cranium, hippocampal progenitor-cell biology is altered. Precursor proliferation is reduced by 62% and neurogenesis is almost entirely ablated after irradiation of the hippocampus. In addition to the defects in the proliferative capacity of the neural progenitor-cell population, the decline in neurogenesis reflects alterations in the microenvironment, including disruption of the microvascular angiogenesis associated with adult neurogenesis and a marked increase in the active microglia within the neurogenic zone. Endogenous stem-cell compartment is not capable of achieving long-lasting repair in the setting of chronic inflammation. The remaining surviving neural precursors proceed as glial cells and fail to differentiate into neurons in the irradiated hippocampus [37,40].
Hippocampal sparing and NCF preservation
Gondi et al. supposed that avoiding the hippocampus during cranial irradiation can preserve NCF [3]. A recent MD Anderson study of low-grade or anaplastic brain tumors treated with radiotherapy observed a dose-response phenomenon, wherein the maximum radiation dose to the left hippocampus was correlated with significant decline in learning and delayed recall [39].
RTOG 0933 is a phase II multi-institutional trial which studied one hundred patients with brain metastases, who received HS-WBRT of dose 30Gy in 10 fractions. Standardized cognitive function and Quality-Of-Life (QOL) assessments were performed at baseline and 2, 4, and 6 months. The primary end point was the Hopkins Verbal Learning Test–Revised Delayed Recall (HVLT-R DR) at 4 months.
Comparison with a historical control of patients treated with WBRT without hippocampal avoidance demonstrated a 30% mean relative decline in HVLT-R DR from baseline to 4 months. The mean relative decline in HVLT-R DR from baseline to 4 months was 7.0% (95% CI, 4.7% to 18.7%), significantly lower in comparison with the historical control. For the 50 patients who were alive at 6 months, HVLT-R DR significantly declined overtime. Higher hippocampal D100% (dose to 100% volume of hippocampus) predicted greater decline in HVLT-R DR. They concluded conformal avoidance of the hippocampus during WBRT is associated with preservation of memory as compared with historical series [41].
Lin et al. studied the impact of HA‑WBRT on NCF preservation in 22 patients with brain metastases. Hippocampal dose parameters were not reported in this study. They suggested that HA‑WBRT preserves critical NCFs in cancer patients harbouring brain metastases with a favorable prognosis [81]. Dios et al. is a recent GICOR-GOECP-SEOR randomized phase III trial of PCI with or without hippocampal avoidance for small-cell lung cancer (PREMER). 150 patients with SCLC randomized to standard PCI (25Gy in 10 fractions) or HA-PCI. Their primary objective was to assess delayed free recall at 3 months and observed comparatively less decline in the HA-PCI arm (5.8%) than the PCI arm (23.5%). Significant decline was also noted in the Total Recall (TR) at 3 months and DFR, TR, and total free recall at 6 months, and TR at 24 months. The incidence of brain metastases, OS, and QOL were not significantly different. They concluded sparing the hippocampus during PCI better preserves cognitive function in patients with SCLC [42].
Apart from anatomical avoidance strategies, the role of neuroprotective agents has been increasingly studied. In RTOG 0614, 554 patients undergoing WBRT for brain metastases with concurrent and adjuvant memantine to placebo were compared. There was a less decline in episodic memory (HVLT-R Delayed Recall) and had significantly longer time to cognitive decline in the memantine arm compared to placebo. The probability of cognitive function failure at 24 weeks was 54% in the memantine arm and 65% in the placebo arm. However, for most cognitive endpoints, no significant differences were observed between memantine and placebo, despite numerical trends favoring the memantine arm [43].
Another trial, CC001, studied cognitive function failure in patients with brain metastases treated by HS-WBRT plus memantine or WBRT plus memantine. HS-WBRT plus memantine preserves cognitive function better but with no difference in progression free survival and OS. They concluded that HS-WBRT should be considered a standard of care for patients with good performance status [44].
Maintaining hippocampal constraints while delivering curative dose of radiation can be challenging. Hofmaier et al. attempted to reduce contralateral hippocampal dose in glioblastoma patients. They were able to reduce contralateral hippocampus generalized equivalent uniform dose with volumetric modulated arc therapy (VMAT) by 36 %, while delivering a total dose of 60Gy in 2Gy per fraction, when compared to the 3D-CRT plans. They demonstrated parietal and a non-temporal tumour localisation as well as a larger target volume predicted a higher hippocampal dose [45].
Kim et al. hypothesized that hippocampal-sparing radiotherapy via VMAT could preserve the NCF of patients with primary brain tumors. They observed verbal memory functions can be preserved by sparing contralateral hippocampus. They achieved a median Dmax of 16.4Gy and a mean dose, expressed as equivalent to a 2-Gy dose (EQD2/2), of 7.4 Gy2 [46].
What can we spare
Sparing the whole hippocampal formation of therapeutic doses of radiation, poses the theoretical risk of intracranial disease progression in these regions. Risk of recurrence or metastases in the avoidance region is largely dependent on the volume of brain tissue that is spared [39,47].
Gondi et al studied the safety profile of RTOG 0933 and did a comprehensive analysis of 371 patients with 1133 brain metastases at presentation and observed brain metastases within 5mm of the hippocampus in 8.6% of patients. 3.0% of 1133 metastasis was located within 5 mm of the hippocampus and none was located within the hippocampus proper [47].
Wee et al suggested hippocampus sparing IMRT (HSRT) does not compromise oncological safety in glioblastoma patients treated to a median dose of 60Gy in 30 fractions. Contralateral hippocampal constraints included Dmax <17Gy. The incidence of failure after 1 year at the contralateral hippocampus and hippocampus + 1 cm was only 2.8% and 6.8% respectively and was mostly seen in patients with disseminated disease after HSRT [48].
Ghia et al. Studied the distribution of brain metastases. In about one hundred patients, they contoured brain metastases as well as hippocampi with 5-, 10-, and 15-mm expansion envelopes in pre–RT MRI T1 weighted images. 3.3% metastases were within 5 mm of the hippocampus and 86.4 % of metastases were greater than 15mm from hippocampus. They concluded that 5-mm margin around the hippocampus for hippocampal sparing WBRT represents an acceptable risk [2].
Therefore, only the dentate gyrus and Cornu Ammonis (CA) fields of the hippocampus proper, which are considered as the sites of adult neurogenesis, are spared. Thus, the avoidance region which is contoured usually include hippocampus proper with a 5mm volumetric expansion [39,47].
Safe dose for the hippocampus
For determining the ideal model of radiation sensitivity, it is essential to ascertain the α/β value of hippocampus. While an α/β value of 2.9 is given for normal brain, the α/β value for hippocampus is still uncertain. Usually, a value of 2 to 3 is used for the hippocampal region but the α/β value for NSCs compartments is 10. Therefore, an α/β ratio of 10 for the true hippocampus and an α/β value of 2 for the whole hippocampus is suggested by some authors. Neurocognitive deficits manifest at doses <10Gy, even as low as 2Gy [49,50].
The recommended maximum dose for the hippocampus (Dmax) is <16Gy and D100% is <9Gy [51, Pokhrel et al, 2016]. Hsu et al. suggested a mean hippocampal dose (normalised to 2Gy fraction) of 5.23Gy2 [Hsypu et al, 2010]. Gondi et al. hypothesised that avoiding the hippocampus during cranial irradiation can preserve NCF and concluded that an EQD2 > 7.3Gy to 40% of the bilateral hippocampus was associated with NCF impairment [3]. However, decreasing the distance of the nearest metastasis from the hippocampus and the total target volume are associated with exceeding hippocampal constraints [54].
Conclusion
The hippocampus is a complex structure which plays a prime role in NCF. The current scientific evidence supports sparing the hippocampus during cranial irradiation can produce clinically significant neurocognitive benefits. These benefits will have more significant clinical translation in young patients and those undergoing curative intent radiotherapy. Therefore, it is necessary that our understanding of dose-dependent radiation-induced NSC dysfunction, the growing knowledge of NSC niches in the brain and development of high precision radiation delivery techniques, should guide us to design and execute NSC-preserving radiotherapy.
Declarations
Conflicts of interest: The authorship no Conflicts of Interest to declare
References
- Brown PD, Gondi V, Pugh S, Tome WA, Wefel JS, Armstrong TS, et al. Hippocampal avoidance during whole-brain radiotherapy plus memantine for patients with brain metastases: phase III trial NRG oncology CC001 [J]. J Clin Oncol. 2020; 38: 1019.
- Ghia A, Tomé WA, Thomas S, Cannon G, Khuntia D, Kuo JS, et al. Distribution of brain metastases in relation to the hippocampus: implications for neurocognitive functional preservation [J]. Int J Radiat Oncol Biol Phys. 2007; 68: 971–7.
- Gondi V, Tolakanahalli R, Mehta MP, Tewatia D, Rowley H, Kuo JS, et al. Hippocampal-sparing whole-brain radiotherapy: a “how-to” technique using helical tomotherapy and linear accelerator–based intensity-modulated radiotherapy [J]. Int J Radiat Oncol Biol Phys. 2010; 78: 1244–52.
- Bir SC, Ambekar S, Kukreja S, Nanda A. Julius Caesar Arantius (Giulio Cesare Aranzi, 1530–1589) and the hippocampus of the human brain: history behind the discovery [J]. J Neurosurg. 2015; 122: 971–5.
- Papez JW. A proposed mechanism of emotion [J]. Arch Neurol Psychiatry. 1937; 38: 725–43.
- MacLean PD. Neurophysiological Symposium: Limbic System and Its Hippocampal Formation. Studies in Animals and Their Possible Application to Man [J]. J Neurosurg. 1954; 11: 29–44.
- Reiner A. An explanation of behavior [J]. Science. 1990; 250: 303–5.
- Brodal A. The hippocampus and the sense of smell; a review [J]. Brain. 1947.
- Douglas RJ. The hippocampus and behavior [J]. Psychol Bull. 1967; 67: 416.
- Kimble DP. Hippocampus and internal inhibition [J]. Psychol Bull. 1968; 70: 285.
- Altman J, Brunner RL, Bayer SA. The hippocampus and behavioral maturation [J]. Behav Biol. 1973; 8: 557–96.
- Scoville WB, Milner B. Loss of recent memory after bilateral hippocampal lesions [J]. J Neurol Neurosurg Psychiatry. 1957; 20: 11.
- O’Keefe J, Nadel L. Précis of O’Keefe & Nadel’s The hippocampus as a cognitive map [J]. Behav Brain Sci. 1979; 2: 487–94.
- O’Keefe J, Dostrovsky J. The hippocampus as a spatial map: preliminary evidence from unit activity in the freely-moving rat [J]. Brain Res. 1971.
- Broadbent NJ, Squire LR, Clark RE. Spatial memory, recognition memory, and the hippocampus [J]. Proc Natl Acad Sci U S A. 2004; 101: 14515–20.
- Andersen P, Morris R, Amaral D, Bliss T, O’Keefe J, editors. The hippocampus book [M]. Oxford: Oxford University Press; 2006.
- Lorente de Nó R. Studies on the structure of the cerebral cortex. II. Continuation of the study of the ammonic system [J]. J für Psychol Neurol. 1934.
- Ramón y Cajal S. Sobre un ganglio especial de la corteza esfeno-occipital [J]. Trab Lab Invest Biol Univ Madrid. 1902; 1: 189–206.
- Witter MP, Doan TP, Jacobsen B, Nilssen ES, Ohara S. Architecture of the entorhinal cortex: a review of entorhinal anatomy in rodents with some comparative notes [J]. Front Syst Neurosci. 2017; 11: 46.
- Swanson LW, Cowan WM. The efferent connections of the suprachiasmatic nucleus of the hypothalamus [J]. J Comp Neurol. 1975; 160: 1–2.
- Felleman DJ, Van Essen DC. Distributed hierarchical processing in the primate cerebral cortex [J]. Cereb Cortex. 1991; 1: 1–47.
- Altman J, Das GD. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats [J]. J Comp Neurol. 1965; 124: 319–35.
- Kaplan MS, Bell DH. Neuronal proliferation in the 9-month-old rodent—radioautographic study of granule cells in the hippocampus [J]. Exp Brain Res. 1983; 52: 1–5.
- Barani IJ, Cuttino LW, Benedict SH, Todor D, Bump EA, Wu Y, et al. Neural stem cell-preserving external-beam radiotherapy of central nervous system malignancies [J]. Int J Radiat Oncol Biol Phys. 2007; 68: 978–85.
- Corkin S. What’s new with the amnesic patient HM? [J]. Nat Rev Neurosci. 2002; 3: 153–60.
- Schacter DL, Wagner AD. Medial temporal lobe activations in fMRI and PET studies of episodic encoding and retrieval [J]. Hippocampus. 1999; 9: 7–24.
- Warrington EK, Weiskrantz L. Amnesic syndrome: consolidation or retrieval? [J]. Nature. 1970; 228: 628–30.
- Pihlajamäki M, Tanila H, Hänninen T, Könönen M, Mikkonen M, Jalkanen V, et al. Encoding of novel picture pairs activates the perirhinal cortex: an fMRI study [J]. Hippocampus. 2003; 13: 67–80.
- Squire LR, Clark RE, Knowlton BJ. Retrograde amnesia [J]. Hippocampus. 2001; 11: 50–5.
- O’Keefe J, Burgess N, Donnett JG, Jeffery KJ, Maguire EA. Place cells, navigational accuracy, and the human hippocampus [J]. Philos Trans R Soc Lond B Biol Sci. 1998; 353: 1333–40.
- Patchell RA. The management of brain metastases [J]. Cancer Treat Rev. 2003; 29: 533–40.
- Armstrong C, Ruffer J, Corn B, DeVries K, Mollman J. Biphasic patterns of memory deficits following moderate-dose partial-brain irradiation: neuropsychologic outcome and proposed mechanisms [J]. J Clin Oncol. 1995; 13: 2263–71.
- Tallet AV, Azria D, Barlesi F, Spano JP, Carpentier AF, Gonçalves A, et al. Neurocognitive function impairment after whole brain radiotherapy for brain metastases: actual assessment [J]. Radiat Oncol. 2012; 7: 1–8.
- Twijnstra A, Boon PJ, Lormans AC, Ten Velde GP. Neurotoxicity of prophylactic cranial irradiation in patients with small cell carcinoma of the lung [J]. Eur J Cancer Clin Oncol. 1987; 23: 983–6.
- Greene-Schloesser D, Moore E, Robbins ME. Molecular pathways: radiation-induced cognitive impairment [J]. Clin Cancer Res. 2013; 19: 2294–300.
- Jalali R, Mallick I, Dutta D, Goswami S, Gupta T, Munshi A, et al. Factors influencing neurocognitive outcomes in young patients with benign and low-grade brain tumors treated with stereotactic conformal radiotherapy [J]. Int J Radiat Oncol Biol Phys. 2010; 77: 974–9.
- Barani IJ, Cuttino LW, Benedict SH, Todor D, Bump EA, Wu Y, et al. Neural stem cell-preserving external-beam radiotherapy of central nervous system malignancies [J]. Int J Radiat Oncol Biol Phys. 2007; 68: 978–85.
- Tofilon PJ, Fike JR. The radioresponse of the central nervous system: a dynamic process [J]. Radiat Res. 2000; 153: 357–70.
- Gondi V, Tomé WA, Mehta MP. Why avoid the hippocampus? A comprehensive review [J]. Radiother Oncol. 2010; 97: 370–6.
- Monje M. Cranial radiation therapy and damage to hippocampal neurogenesis [J]. Dev Disabil Res Rev. 2008; 14: 238–42.
- Gondi V, Mehta MP, Pugh S, Tome WA, Kanner A, Caine C, et al. Memory preservation with conformal avoidance of the hippocampus during whole-brain radiation therapy for patients with brain metastases: primary endpoint results of RTOG 0933. Int J Radiat Oncol Biol Phys. 2013; 87: 1186.
- Rodríguez de Dios N, Couñago F, Murcia-Mejía M, Rico-Oses M, Calvo-Crespo P, Samper P, et al. Randomized phase III trial of prophylactic cranial irradiation with or without hippocampal avoidance for small-cell lung cancer (PREMER): a GICOR-GOECP-SEOR study. J Clin Oncol. 2021; 39: 3118–27.
- Brown PD, Pugh S, Laack NN, Wefel JS, Khuntia D, Meyers C, et al. Memantine for the prevention of cognitive dysfunction in patients receiving whole-brain radiotherapy: a randomized, double-blind, placebo-controlled trial. Neuro Oncol. 2013; 15: 1429–37.
- Brown PD, Gondi V, Pugh S, Tome WA, Wefel JS, Armstrong TS, et al. Hippocampal avoidance during whole-brain radiotherapy plus memantine for patients with brain metastases: phase III trial NRG oncology CC001. J Clin Oncol. 2020; 38: 1019.
- Hofmaier J, Kantz S, Söhn M, Dohm OS, Bächle S, Alber M, et al. Hippocampal sparing radiotherapy for glioblastoma patients: a planning study using volumetric modulated arc therapy. Radiat Oncol. 2016; 11: 1–8.
- Kim KS, Wee CW, Seok JY, Hong JW, Chung JB, Eom KY, et al. Hippocampus-sparing radiotherapy using volumetric modulated arc therapy (VMAT) to the primary brain tumor: the result of dosimetric study and neurocognitive function assessment. Radiat Oncol. 2018; 13: 1–8.
- Gondi V, Tome WA, Marsh J, Struck A, Ghia A, Turian JV, et al. Estimated risk of perihippocampal disease progression after hippocampal avoidance during whole-brain radiotherapy: safety profile for RTOG 0933. Radiother Oncol. 2010; 95: 327–31.
- Wee CW, Kim KS, Kim CY, Han JH, Kim YJ, Kim IA. Feasibility of hippocampus-sparing VMAT for newly diagnosed glioblastoma treated by chemoradiation: pattern of failure analysis. Radiat Oncol. 2020; 15: 1–8.
- Hall EJ, Giacci AJ. Dose–response relationships for model normal tissues. In: Hall EJ, Giacci AJ, editors. Radiobiology for the radiologist. 6th ed. Philadelphia: Lippincott Williams & Wilkins; 2006. p. 303–26.
- Kazda T, Jancalek R, Pospisil P, Sevela O, Prochazka T, Vrzal M, et al. Why and how to spare the hippocampus during brain radiotherapy: the developing role of hippocampal avoidance in cranial radiotherapy. Radiat Oncol. 2014; 9: 1–10.
- Wan JF, Zhang SJ, Wang L, Zhao KL. Implications for preserving neural stem cells in whole brain radiotherapy and prophylactic cranial irradiation: a review of 2270 metastases in 488 patients. J Radiat Res. 2013; 54: 285–91.
- Pokhrel D, Sood S, McClinton C, Shen X, Lominska C, Saleh H, et al. Treatment planning strategy for whole-brain radiotherapy with hippocampal sparing and simultaneous integrated boost for multiple brain metastases using intensity-modulated arc therapy. Med Dosim. 2016; 41: 315–22.
- Hsu F, Carolan H, Nichol A, Cao F, Nuraney N, Lee R, et al. Whole brain radiotherapy with hippocampal avoidance and simultaneous integrated boost for 1–3 brain metastases: a feasibility study using volumetric modulated arc therapy. Int J Radiat Oncol Biol Phys. 2010; 76: 1480–5.
- Birer SR, Olson AC, Adamson J, Hood R, Susen M, Kim G, et al. Hippocampal dose from stereotactic radiosurgery for 4 to 10 brain metastases: risk factors, feasibility of dose reduction via re-optimization, and patient outcomes. Med Dosim. 2017; 42: 310–16.